|
|
Short Communication
Survival of honey bees (Apis mellifera) infected with Crithidia mellificae spheroid forms (Langridge and McGhee: ATCC ® 30254 ™) in the presence of Nosema ceranae
Mariano Higes
Junta de Comunidades de Castilla-La Mancha, IRIAF, Centro de Investigación Apícola y Agroambiental (CIAPA), Laboratorio de
Patología Apícola, 19180 Marchamalo (Guadalajara). Spain
Cristina Rodríguez-García
Junta de Comunidades de Castilla-La Mancha, IRIAF, Centro de Investigación Apícola y Agroambiental (CIAPA), Laboratorio de
Patología Apícola, 19180 Marchamalo (Guadalajara). Spain
Tamara Gómez-Moracho
Junta de Comunidades de Castilla-La Mancha, IRIAF, Centro de Investigación Apícola y Agroambiental (CIAPA), Laboratorio de
Patología Apícola, 19180 Marchamalo (Guadalajara). Spain
Aranzazu Meana
Universidad Complutense de Madrid, Facultad de Veterinaria, Departamento de Sanidad Animal, 28040 Madrid. Spain
Carolina Bartolomé
Universidade de Santiago de Compostela, CIMUS, Medicina Xenómica, 15782 Santiago de Compostela. Spain
IDIS, Xenómica Comparada de Parásitos Humanos, Santiago de Compostela. Spain
Xulio Maside
Universidade de Santiago de Compostela, CIMUS, Medicina Xenómica, 15782 Santiago de Compostela. Spain
IDIS, Xenómica Comparada de Parásitos Humanos, Santiago de Compostela. Spain
Universidade de Santiago de Compostela, Departamento de Anatomía Patolóxica e Ciencias Forenses, 15782 Santiago de Compostela.
Spain
Laura Barrios
Consejo Superior Investigaciones Científicas, SGAI. Departamento de Estadística, 28006 Madrid. Spain
Raquel Martín-Hernández
Junta de Comunidades de Castilla-La Mancha, IRIAF, Centro de Investigación Apícola y Agroambiental (CIAPA), Laboratorio de
Patología Apícola, 19180 Marchamalo (Guadalajara). Spain
Fundación Parque Científico y Tecnológico de Albacete, Instituto de Recursos Humanos para la Ciencia y la Tecnología (INCRECYT),
Albacete, Spain
| |
Abstract
Crithidia mellificae, a trypanosomatid parasite of Apis mellifera, has been proposed to be one of the pathogens responsible for the serious honey bee colony losses produced worldwide in the
last decade, either alone or in association with Nosema ceranae. Since this pathogenic effect contradicts the results of the experimental infections originally performed by Langridge and
McGhee nearly 40 years ago, we investigated the potential linkage of this protozoan with colony decline under laboratory conditions.
Nosema-free and trypanosomatid-free honey bees from three different colonies were experimentally infected with fresh C. mellificae spheroid forms (reference strain ATCC30254), with N. ceranae fresh spores and with both parasites at the same time. Replicate cages were kept at 27 °C and used to analyse survival. C. mellificae spheroid forms did not reduce significantlythe survival of the worker bees (64.5% at 30 days post-infection vs. 77.8% for the uninfected bees used as controls;differences were non statistically significant) under these experimental conditions. In contrast, the cages infected with N. ceranae exhibited higher rates of mortality from the 20th day post-infection onwards, irrespective of the presence of C. mellificae, suggesting that the spheroid forms of the latter have no pathological effect on A. mellifera.
Additional key words:
trypanosomatids; microsporidia; colony collapse.
Abbreviations used:
CCD (colony collapse disorder); dpi (days post-infection); PCR (polymerase chain reaction).
Authors’ contributions: Conceived and designed the experiments: MH, RMH, AM and CB. Performed the experiments: MH, CRG, TGM and RMH. Analyzed the
data: MH, RMH,AM, CB, XM and LB. Statistical analysis: LB. Coordinating the research project: MH. Wrote the paper: MH, RMH,
XM and CB.
Citation: Higes, M.; Rodriguez-García, C.; Gómez-Moracho, T.; Meana, A.; Bartolomé, C.; Maside, X.; Barrios, L.; Martín-Hernández, R.
(2016). Short communication: Survival of honey bees (Apis mellifera) infected with Crithidia mellificae (Langridge and McGhee: ATCC® 30254™) in the presence of Nosema ceranae. Spanish Journal of Agricultural Research, Volume 14, Issue 3, e05SC02. http://dx.doi.org/10.5424/sjar/2016143-8722.
Received: 30 Sep 2015. Accepted: 29 Jun 2016.
Copyright © 2016 INIA. This is an open access article distributed under the terms of the Creative Commons Attribution-Non Commercial (by-nc) Spain
3.0 Licence, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly
cited.
Funding: INIA-FEDER (RTA2013-00042-C10-06 and E-RTA2014-00003-C03).
Competing interests: None of the authors of this paper has any financial or personal relationship with other individuals or organizations that
could have inappropriately influenced or bias the content of the paper. The authors declare that they have no competing interests.
Correspondence should be addressed to Mariano Higes: mhiges@jccm.es.
|
IntroductionTop
In the last decade, the phenomenon of honey bee colony death has attracted the interest of scientists, beekeepers and the
general public alike. Increased annual honey bee colony losses have been associated with many factors, including viruses,
bacteria, fungi, mites and trypanosomatids (Higes et al., 2006; vanEngelsdorp et al., 2009; Cox-Foster et al., 2007; Dainat et al., 2012; Francis et al., 2013; Ravoet et al., 2013; Cepero et al., 2014). Trypanosomatids are flagellated protozoan parasites and they are the direct descendants of the first eukaryotes that appeared
on the earth ~1.5 billion years ago (Knoll, 1992). Several genera of the Trypanosomatidae cause disease in humans, domestic animals, plants (Galanti et al., 1998) and insects (Schmid-Hempel & Tognazzo, 2010). Indeed, the interest in insect trypanosomatids, especially those affecting bees, has increased among researchers due to
their possible involvement in colony collapse disorder (CCD). Until now honey bees were thought to be infected by only a single
trypanosomatid species, Crithidia mellificae, the trypanosomatid first detected in Australia during the diagnostic examination of Apis mellifera apiaries (Langridge & McGhee, 1967). The original experimental infection with C. mellificae suggested it had no pathological effects on this hymenopteran host, which explains why this organism has been largely ignored
for more than 40 years.
However, a relationship between C. mellificae infection and colony losses was recently proposed in the USA (Runckel et al., 2011) and Belgium (Ravoet et al., 2013). Indeed, C. mellificae was 6.15-fold more abundant in CCD-affected colonies than in unaffected colonies in a colony-cohort study, and its detection
was strongly associated with the presence of N. ceranae (Runckel et al., 2011). In addition, a high prevalence of C. mellificae (70.5%) was reported in Belgium (Ravoet et al., 2013), and a correlation was detected between C. mellificae incidence in July and winter mortality. This study also confirmed the link between C. mellificae and N. ceranae previously described in the USA (Runckel et al., 2011), suggesting that co-infection by these pathogens has a negative synergistic impact on colony longevity (Ravoet et al., 2013). More recently, a holistic screening of professional apiaries with typical honey bee depopulation symptoms in Spain (Cepero et al., 2014) revealed a higher prevalence of N. ceranae with only a moderate presence of trypanosomatids. Genetic analyses of the 18S rDNA and GAPDH sequences of these trypanosomatids
proved that they were phylogenetically distant from those of the reference ATCC30254 C. mellificae strain, providing the first hard evidence of honey bee infection by a different lineage (Cepero et al., 2014). The existence of this taxon, which was later named Lotmaria passim (Schwarz et al., 2015), had been previously suggested in the USA (Cornman et al., 2012). It is also worth noting that all the sequences attributed to C. mellificae until then, including its draft genome assembly (Runckel et al., 2014), were more closely related to this new taxon than to the “true” C. mellificae (Cepero et al., 2014).
Together, these studies inspired renewed interest in this pathogen and its relationship with the highly prevalent microsporidium
N. ceranae as one of the possible causes of bee colony collapse in different parts of the world. Given the discrepancies regarding the
pathological effect of C. mellificae in honey bees (Langridge & McGhee, 1967; Ravoet et al., 2013), we evaluated the effect of experimentally infecting bee colonies under laboratory conditions with the C. mellificae ATCC30254 reference strain, with or without N. ceranae. In this way we were able to specifically assess the influence of C. mellificae on the bees’ life expectancy and determine whether a synergistic effect of this pathogen with N. ceranae could be detected.
Material and methodsTop
To perform these experiments, the C. mellificae reference strain ATCC30254 was first cultured in ATCC medium 355 as recommended, and it was further sub-cultured in solid
medium as described by Popp & Lattorff (2011). Visible, isolated colonies were selected and resuspended in milliQ water (PCR-quality) to obtain trypanosomatid spheroid
forms for experimental infection (Schwarz & Evans, 2013). The DNA from those colonies was extracted as described previously in Cepero et al. (2014) and the trypanosomatid species was confirmed by PCR (Meeus et al., 2010) and subsequent sequencing (Cepero et al., 2014). Freshly purified N. ceranae spores for experimental infection were obtained as described elsewhere (Martín-Hernández et al., 2011, 2012; Higes et al., 2013), counting both the N. ceranae spores and C. mellificae spheroid forms in a haemocytometer. DNA was extracted from an aliquot of Nosema spores to confirm the species by PCR (Martín-Hernández et al., 2012). Nosema-free and trypanosomatid-free honey bees were obtained as described previously (Higes et al., 2013). Emerging worker bees from three different colonies were removed carefully, confined to cages in groups of 25 and kept in
the incubator for 5 days at 27ºC, the optimum temperature for C. mellificae growth (Langridge & McGhee, 1967). Sixteen replicate cages were prepared, 12 of which were used to analyse survival: three cages infected with fresh N. ceranae spores; three with fresh C. mellificae spheroid forms; three co-infected with N. ceranae and C. mellificae; and three that remained uninfected (used as controls).
Honey bees (Apis mellifera iberiensis) were infected as described elsewhere (Higes et al., 2013), administering 1 µL of a sucrose solution containing 50,000 N. ceranae spores or 50,000 C. mellificae spheroid forms. The co-infected bees received 1 µL of each pathogen solution, while uninfected control bees were fed with
2 µL of the 50% sucrose solution alone. After inducing infection, the bees in each cage were fed ad libitum with 50% sugar
syrup + 2% Promotor L (Calier Lab). Infected and uninfected bees were kept at 27 °C in separate incubators (Memmert ® Mod.
IPP500, 0.1 °C), and the replicate cages were observed daily. Cumulative honey bee mortality was analysed using a Kaplan-Meier
Log-Rank survival analysis and the slope of the survival function was analysed according to the Mantel-Cox test with the aid
of IBM Statistics V22.0.
Results and discussionTop
The mortality of the workers infected with N. ceranaeand N. ceranae+C. mellificae was significantly higher than that of those that received the C. mellificae treatment alone and that of the control worker bees. Bee survival at 30 days post-infection (dpi) was 77.8%, 64.5%, 0% and
0% for the uninfected control honey bees, and the honey bees from cages infected with C. mellificae, N. ceranae and a mixture of both parasites, respectively (Fig. 1). The mortality of the workers infected with N. ceranae and N. ceranae+C. mellificae was significantly higher than that of those that received C. mellificae alone and that of the control worker bees (Kaplan-Meier Long-Rank test, p<0.001). Conversely, the survival of the control and the C. mellificae infected bees was significantly higher than that of the other two groups (Kaplan-Meier Long-Rank test, p<0.001). The slopes of the survival functions were statistically different according to the Mantel-Cox test (p<0.05). The evolution of control worker bee survival was similar and not significantly different from that of those infected
with C. mellificae alone (p>0.05). Likewise, the N. ceranae and mixed infection groups also displayed a parallel evolution in terms of survival (p>0.05), which differed considerably from the other two groups from 20 dpi onwards. The highest mortality rates of worker honey
bees occurred in the cages infected with N. ceranae, irrespective of the presence of C. mellificae.
|
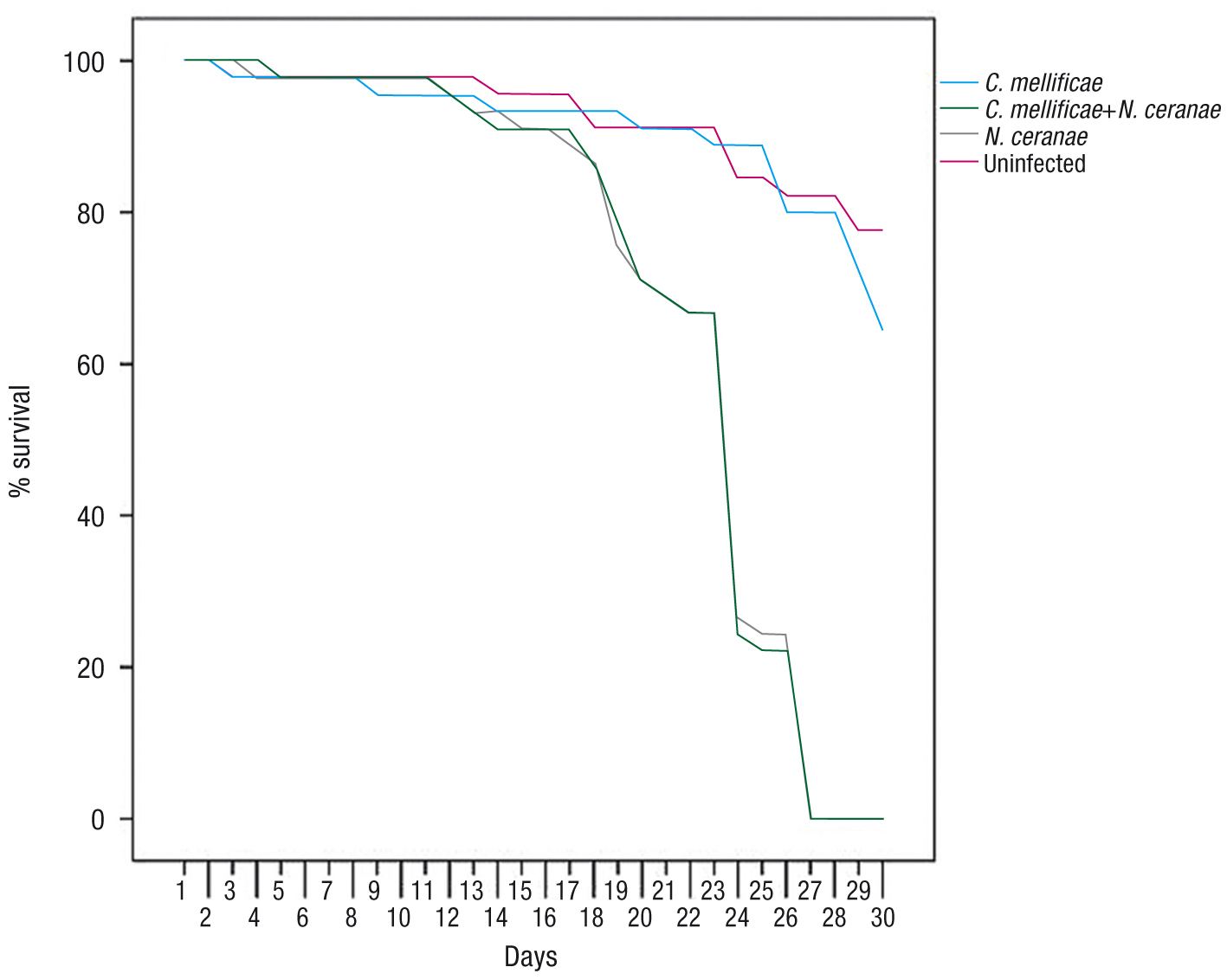 Figure 1. Cumulative honey bee mortality after infection with Crithidia mellificae, Nosema ceranae and N. ceranae+C. mellificae or uninfected (Control).
Figure 1. Cumulative honey bee mortality after infection with Crithidia mellificae, Nosema ceranae and N. ceranae+C. mellificae or uninfected (Control).
|
Since the first report of N. ceranae infecting of A. mellifera bees there has been some controversy about the effects of these microsporidia. However, most of the studies published in
recent years have confirmed that N. ceranae has a pathogenic effect on this host, expressed as a shortening of the honey bee (workers) lifespan (Higes et al., 2013). Significantly, the longevity of infected bees varies strongly among these studies, probably due to the many factors that
influence the development and the course of the infection. For example, this is the case of the temperature, which must be
taken into consideration in laboratory studies since N. ceranae has been proposed to be eurithermal (Higes et al., 2010). In this study, the survival of honey bees following experimental infection with N. ceranae is consistent with that seen previously (reviewed by Higes et al., 2013; Williams et al., 2014; Huang et al., 2015; Milbrath et al., 2015), confirming that bees infected with N. ceranae have a shorter life expectancy than uninfected bees. The incubation temperature used may explain the distinct evolution in
the survival of infected honey bees relative to other laboratory experiments performed in our laboratory (Martín-Hernandez et al., 2009).
Previous co-infection experiments with N. ceranae and C. mellificae under laboratory conditions have shown to modify the bees’s immune response in a distinct manner to that produced when bees
were exposed only to one pathogen or another. Mixed C. mellificae and N. ceranae infection altered the repertoire of systemic antimicrobial peptides and reduced cellular immunity at the level of the immune
response, although the mortality of the infected bees was not recorded (Schwarz & Evans, 2013). An epidemiological link between the presence of C. mellificae and bee colony collapse has been suggested in field conditions, either alone or in association with N. ceranae (Runckel et al., 2011; Ravoet et al., 2013), although no information regarding worker bee mortality was reported. The effect of the infection with C. mellificae spheroid forms on honey bee survival suggests that such cellular stage does not alter the insect’s longevity, and does not
significantly modify the mortality of bees co-infected with N. ceranae when compared to that of the bees infected with the microsporidium alone. These results are consistent with those of Langridge & McGhee (1967) suggesting that C. mellificae has no pathological effect on honey bees. However, further research is needed to determine the development of this trypanosomatid
in different culture media, investigate its life cycle and the possible variability of its pathogenic capacity. The finding
of a new trypanosomatid taxon in collapsed colonies under field conditions raises new questions about the role of C. mellificae in honey bee losses, an issue that should be addressed through appropriate experimental infection and epidemiological analyses.
AcknowledgementsTop
The authors wish to thank J. Almagro, J. García, V. Albendea, C. Uceta, M. Gajero and T. Corrales at Laboratorio de Patología
Apícola, Centro de Investigación Apícola y Agroambiental (CIAPA), IRIAF, Junta de Comunidades de Castilla-La Mancha for their
technical support. We also thank Prof. de Graaf and Dr. Ravoet for supplying the ATCC30254 reference strain and for sharing
their knowledge about C. mellificae.
ReferencesTop
|
|
|
○
|
Cepero A, Gómez-Moracho T, Bernal JL, Del Nozal MJ, Bartolomé C, Maside X, Meana A, González-Porto AV, de Graaf DC, Martín-Hernández
R, Higes M, 2014. Holistic screening of collapsing honey bee colonies in Spain: a case study. BMC Research Notes 7: 649. http://dx.doi.org/10.1186/1756-0500-7-649.
|
|
○
|
Cornman RS, Tarpy DR, Chen Y, Jeffreys R, Lopez D, Pettis JS, vanEngelsdorp D, Evans JD, 2012. Pathogen webs in collapsing
honey bee colonies. PLOS ONE 8: e43652. http://dx.doi.org/10.1371/journal.pone.0043562.
|
|
○
|
Cox-Foster DL, Conlan S, Holmes E., Palacios E, Evans JD, et al., 2007. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 318: 283-287. http://dx.doi.org/10.1126/science.1146498.
|
|
○
|
Dainat B, Evans JD, Chen YP, Gauthier L, Neumann P, 2012. Predictive markers of honey bee colony collapse. PLOS ONE 7(2):
e32151. http://dx.doi.org/10.1371/journal.pone.0032151.
|
|
○
|
Francis RM, Nielsen SL, Kryger P, 2013. Varroa-virus interaction in collapsing honey bee colonies. PLOS ONE 8(3):e57540. http://dx.doi.org/10.1371/journal.pone.0057540.
|
|
○
|
Galanti N, Galindo M, Sabaj V, Espinoza I, Toro G, 1998. Histone genes in trypanosomatids. Parasitol Today 14: 64-70. http://dx.doi.org/10.1016/S0169-4758(97)01162-9.
|
|
○
|
Higes M, Martín-Hernández R, Meana A, 2006. Nosema ceranae, a new microsporidian parasite in honeybees in Europe. J Invertebr Pathol 92: 93-95. http://dx.doi.org/10.1016/j.jip.2006.02.005.
|
|
○
|
Higes M, García-Palencia P, Botías C, Meana A, Martín-Hernández R, 2010. The differential development of microsporidia infecting
worker honey bee (Apis mellifera) at increasing incubation temperature. Environ Microbiol Rep 2: 745-748. http://dx.doi.org/10.1111/j.1758-2229.2010.00170.x.
|
|
○
|
Higes M, Juarranz Á, Dias-Almeida J, Lucena S, Botías C, Meana A, Garcia-Palencia P, Martín-Hernández R, 2013. Apoptosis in
the pathogenesis of Nosema ceranae (Microsporidia: Nosematidae) in honey bees (Apis mellifera). Environ Microbiol Rep 5(4): 530-536. http://dx.doi.org/10.1111/1758-2229.12059.
|
|
○
|
Huang WF, Solter L, Aronstein K, Huang Z, 2015. Infectivity and virulence of Nosema ceranae and Nosema apis in commercially available North American honey bees. Invertebr Pathol 124:107-113. http://dx.doi.org/10.1016/j.jip.2014.10.006.
|
|
○
|
Knoll AJ, 1992. The early evolution of eukaryotes: a geological perspective. Science 256: 622-627. http://dx.doi.org/10.1126/science.1585174.
|
|
○
|
Langridge D, McGhee R, 1967. Crithidia mellificae n. sp. An acidophilic trypanosomatid of de honey bee Apis mellifera. J Protozool 14(3): 485-487. http://dx.doi.org/10.1111/j.1550-7408.1967.tb02033.x.
|
|
○
|
Martín-Hernández R, Meana A, García-Palencia P, Marín P, Botías C, Garrido-Bailó E, Barrios L, Higes M, 2009. Effect of temperature
on the biotic potential of honeybee microsporidia. Appl Environ Microbiol 75 (8): 2554-2557. http://dx.doi.org/10.1128/AEM.02908-08.
|
|
○
|
Martín-Hernández R, Botías C, Barrios L, Martínez-Salvador A, Meana A, Mayack C, Higes M, 2011. Comparison of the energetic
stress associated with experimental Nosema ceranae and Nosema apis infection of honey bees (Apis mellifera). Parasitol Res 109: 605-612. http://dx.doi.org/10.1007/s00436-011-2292-9.
|
|
○
|
Martín-Hernández R, Botías C, Garrido Bailón E, Martínez-Salvador A, Prieto L, Meana A, Higes M, 2012. Microsporidia infecting
Apis mellifera: coexistence or competition. Is Nosema ceranae replacing Nosema apis? Environ Microbiol 14 (8): 2127-2138. http://dx.doi.org/10.1111/j.1462-2920.2011.02645.x.
|
|
○
|
Meeus I, De Graaf DC, Jans K, Smagghe G, 2010. Multiplex PCR detection of slowly-evolving trypanosomatids and neogregarines
in bumblebees using broad-range primers. J Appl Microbiol 109: 107-115.
|
|
○
|
Milbrath MO, van Tran T, Huang W-F, Solter, LF, Tarpy DR, Lawrence F, Huang Z, 2015. Comparative virulence and competition between Nosema apis and Nosema ceranae in honey bees (Apis mellifera). J Invertebr Pathol 125: 9-15. http://dx.doi.org/10.1016/j.jip.2014.12.006.
|
|
○
|
Popp M, Lattorff HMG, 2011. A quantitative in vitro cultivation technique to determine cell number and growth rates in strains
of Crithidia bombi (Trypanosomatidae), a parasite of bumblebees. J Eukaryotic Microbiol 58: 7-10. http://dx.doi.org/10.1111/j.1550-7408.2010.00514.x.
|
|
○
|
Ravoet J, Maharramov J. Meeus I, De Smet L, Wenseleers T, Smagghe G, de Graaf DC, 2013. Comprehensive bee pathogen screening
in Belgium reveals Crithidia mellificae as a new contributory factor to winter mortality. PLOS ONE 8: e72443. http://dx.doi.org/10.1371/journal.pone.0072443.
|
|
○
|
Ravoet J, Schwarz RS, Descamps T, Yañez O, Torkaz OC, Martín-Hermández R, Bartolomé C, De Smert L, Higes M, Wenseleers T,
Schmid-Hempel R, Neuman P, Kadowaki T, Evans JD, de Graaf D, 2015. Differential diagnosis of the honey bee trypanosomatids
Crithidia mellificae and Lotmaria passim. J Invertebr Pathol 130: 21-27. http://dx.doi.org/10.1016/j.jip.2015.06.007.
|
|
○
|
Runckel C, Flenniken M, Engel JC, Ruby JG, Ganem D, Andino R, DeRisi JL, 2011. Temporal analysis of the honey bee microbiome
reveals four novel viruses and seasonal prevalence of known viruses, Nosema, and Crithidia. PLoS ONE 6: e20656. http://dx.doi.org/10.1371/journal.pone.0020656.
|
|
○
|
Runckel C, DeRisi J, Flenniken ML, 2014. A draft genome of the honey bee trypanosomatid parasite Crithidia mellificae. PLOS ONE 9: e95057. http://dx.doi.org/10.1371/journal.pone.0095057.
|
|
○
|
Schmid-Hempel R, Tognazzo M, 2010. Molecular divergence defines two distinct lineages of Crithidia bombi (Trypanosomatidae), parasites of bumblebees. J Eukaryotic Microbiol 57 (4): 337-345. http://dx.doi.org/10.1111/j.1550-7408.2010.00480.x.
|
|
○
|
Schwarz RS, Evans JD, 2013. Single and mixed-species trypanosome and microsporidia infections elicit distinct, ephemeral cellular
and humoral immune responses in honey bees. Dev Comp Immunol 40 (3-4): 300-310. http://dx.doi.org/10.1016/j.dci.2013.03.010.
|
|
○
|
Schwarz RS, Bauchan G., Murphy CA, Ravoet J, De Graaf DC. Evans JD, 2015. Characterization of two species of trypanosomatidae
from the honey bee Apis mellifera: Crithidia mellificae Langridge and McGhee, 1967 and Lotmaria passim n. gen., n. sp. J Eukaryotic Microbiol 10: 1-17.
|
|
○
|
Vanengelsdorp D, Evans JD, Saegerman C, Mullin C, Haubruge E, Nguyen BK, Frazier M, Frazier J, Cox-Foxter D, Chen Y et al., 2009. Colony collapse disorder: a descriptive study. PLoS ONE 4: e6481. http://dx.doi.org/10.1371/journal.pone.0006481.
|
|
○
|
Williams GR, Shutler D, Burgher-MacLellan K., Rogers RE, 2014. Infra-population and community dynamics of the parasites Nosema apis and Nosema ceranae, and consequences for honey bee (Apis mellifera) hosts. PLOS ONE 9: e99465. http://dx.doi.org/10.1371/journal.pone.0099465.
|